Роль вкусовой чувствительности. Вкусовая чувствительность Вкусовая чувствительность
1. Вестибулярная сенсорная система. Функция вестибулярной системы. Вестибулярный аппарат. Костный лабиринт. Перепончатый лабиринт. Отолиты.
2. Волосковые клетки. Свойства рецепторных клеток вестибулярного аппарата. Стереоцилии. Киноцилий.
3. Отолитовый аппарат. Отолитовый орган. Адекватные раздражители рецепторов отолитовых органов.
4. Полукружные каналы. Адекватные раздражители рецепторов полукружных каналов.
5. Центральная часть вестибулярной системы. Вестибулярные ядра. Кинетозы.
6. Вкус. Вкусовая чувствительность. Вкусовая сенсорная система. Вкусовая рецепция. Вкусовая пора.
8. Центральный отдел вкусовой системы. Проводящие пути вкусовой чувствительности. Ядра вкуса.
9. Вкусовое восприятие. Обонятельная сенсорная система. Макросматики. Микросматики.
10. Запах (запахи). Классификация запахов. Стереохимическая теория запахов.
Мембрана микроворсинок вкусовых клеток содержит специфические участки (рецепторы), предназначенные для связывания растворенных в жидкой среде полости рта химических молекул. Существует четыре разновидности вкусовых ощущений, или четыре вкусовые модальности: сладкое, кислое, соленое и горькое. Строгой зависимости между химической природой вещества и вкусовым ощущением нет : например, сладким вкусом обладают не только сахара, но и некоторые неорганические соединения (соли свинца, бериллия), а самым сладким веществом является не усваиваемый организмом сахарин. Большинство вкусовых клеток полимодальны, т. е. могут реагировать на стимулы всех четырех вкусовых модальностей.
Присоединение к специфическим рецепторам молекул, обладающих сладким вкусом, активирует систему вторичных посредников аденилатциклазы - циклического аденозинмонофосфата, которые закрывают мембранные каналы ионов калия, и поэтому мембрана рецепторной клетки деполяризуется. Вещества, обладающие горьким вкусом, активируют одну из двух систем вторичных посредников: 1) фосфолипазу С - инозитол-3-фос-фат, что приводит к выходу из внутриклеточного депо ионов кальция с последующим выделением медиатора из рецепторной клетки; 2) специфический G-белок гастдуцин, регулирующий внутриклеточную концентрацию цАМФ, которая управляет катионными каналами мембраны и этим определяет возникновение рецепторного потенциала. Действие на рецепторы молекул, имеющих соленый вкус, сопровождается открытием управляемых натриевых каналов и деполяризацией вкусовой клетки. Вещества, обладающие кислым вкусом, закрывают мембранные каналы для ионов калия, что ведет к деполяризации рецепторной клетки.
Величина рецепторного потенциала зависит от вкусового качества и концентрации химического вещества , действующего на клетку. Возникновение рецепторного потенциала приводит к выделению вкусовой клеткой медиатора, действующего через синапс на афферентное волокно первичного сенсорного нейрона, в котором через 40-50 мс от начала действия стимула повышается частота потенциалов действия. Возникшие в афферентных волокнах нервные импульсы проводятся к ядрам одиночных пучков продолговатого мозга. При повышении концентрации действующего вещества общее количество реагирующих чувствительных волокон возрастает за счет вовлечения высокопороговых афферентов в передачу информации от рецепторов.
Вкусовая чувствительность
Пороги вкусовой чувствительности выявляются поочередным нанесением на поверхность языка растворов веществ, обладающих разными вкусовыми качествами (табл. 17.4). Абсолютным порогом чувствительности считают появление определенного вкусового ощущения, отличающегося от вкуса дистиллированной воды. Вкус одного и того же вещества может восприниматься по-разному в зависимости от его концентрации в растворе; например, при малой концентрации хлорида натрия он ощущается сладким, а при большей концентрации - соленым. Максимальная способность различать концентрацию растворов одного и того же вещества и, соответственно, самый низкий дифференциальный порог вкусовой чувствительности характерны для среднего диапазона концентраций, а при высоких концентрациях вещества дифференциальный порог повышается.
Таблица 17.4. Абсолютные пороги восприятия веществ с характерным вкусом
Абсолютные пороги вкусовой чувствительности индивидуально различаются, но у подавляющего большинства людей самым низким оказывается порог определения веществ с горьким вкусом. Эта особенность восприятия возникла в эволюции, она способствует отказу от употребления в пищу веществ горького вкуса, к которым принадлежат алкалоиды многих ядовитых растений. Вкусовые пороги различаются у одного и того же человека в зависимости от его потребности в тех или иных веществах, они повышаются вследствие длительного употребления веществ с характерным вкусом (например, сладостей или соленостей) или курения, потребления алкоголя, обжигающих напитков. Разные области языка различаются вкусовой чувствительностью к различным веществам, что обусловлено особенностями распределения вкусовых рецепторов. Кончик языка более других областей чувствителен к сладкому, боковые стороны языка - к кислому и соленому, а корень языка - к горькому. Вкусовые ощущения в большинстве случаев мультимодальны и основаны не только на избирательной химической чувствительности вкусовых рецепторных клеток, но и на раздражении пищей терморецепторов и механорецепторов ротовой полости , а также действии летучих компонентов пищи на обонятельные рецепторы .
ВКУС И ОБОНЯНИЕ
X. Альтнер, И. Бекх
13.1. Характеристика химических ощущений
Ощущения вкуса и запаха обусловлены избирательной и высокочувствительной реакцией специализированных сенсорных клеток на присутствие молекул определенных соединений. В более широком смысле специфические реакции на химические вещества, например гормоныили нейромедиаторы, характерны для многих клеток и тканей. Однако вкусовые и обонятельные сенсорные клетки действуют как экстероцепторы; их реакции дают важную информацию о внешних стимулах, обрабатываемую особыми участками мозга, которые и отвечают за соответствующие ощущения. Другие хеморецепторные клетки служат интероцепторами, определяющими, например, уровень СО 2 (разд. 21.6).
Вкус и запах можно охарактеризовать и различить на основании морфологических и физиологических критериев. Различия между двумя этими видами ощущений наиболее очевидны при сравнении типов (качеств) адекватных для них стимулов (табл. 13.1). Другие характеристики, например чувствительность к стимулам или физическое состояние последних, хотя в целом и неодинаковы, но могут и взаимоперекрываться.
По сравнению с другими ощущениями у вкуса и обоняния значительно выше адаптируемость (ср. рис. 8.5). При длительном воздействии стимула возбуждение в афферентных путях заметно ослабляется, соответственно ослабляется и восприятие; например, уже очень быстро в среде даже с сильным запахом мы перестаем его ощущать. В равной мере характерна для химических ощущений и высокая чувствительность к определенным стимулам. Вместе с тем диапазон различаемых интенсивностей стимуляции сравнительно невелик (1:500), а порог различения высок. Показатель в степенной функции Стивенса ψ = k (φ - φо) a равен 0,4–0,6 для запаха и около 1–для вкусовых стимулов (ср. рис. 8.14).
Первичные процессы и химическая специфичность .
Первое событие при стимуляции хеморецепторов это, согласно современным представлениям, химическое взаимодействие, основанное на слабом связывании адекватной молекулы с рецепторным белком. Из вкусовых органов выделены белки с ферментативными свойствами, субстратная
специфичность и кинетические особенности которых такие же, как у самих рецепторов. Последующие события, приводящие к электрической реакции клеточной мембраны, неизвестны. Каждая рецепторная клетка высокоизбирательно реагирует на специфическую группу веществ. Малейшие изменения в структуре молекулы могут изменить характер ее восприятия или сделать ее неадекватным стимулом. Стимулирующая эффективность соединения, вероятно, наиболее существенно зависит от его размера (например, длины цепи) и внутреннего распределения электрических зарядов (т. е. расположения функциональных групп). Однако тот факт, что во многих случаях молекулы веществ, сильно различающихся по химическому строению, вызывают одинаковые обонятельные ощущения, пока не получил объяснения. Например, три приведенных ниже вещества, несмотря на их структурные различия, обладают одинаковым мускусным запахом (см. Beets в ).
Высказано предположение, что хеморецепторы содержат рецепторные центры, специфичные в отношении определенных групп веществ. Такая точка зрения подтверждается случаями частичной аносмии, т. е. нечувствительности к запаху некоторых, очень близких химических соединений. Сходным образом можно интерпретировать избирательное действие некоторых лекарственных препаратов на орган вкуса. Нанесение на язык гимнемата калия (вещества, выделенного из индийского растения Gymnema silvestre ) приводит к утрате только восприятия сладкого–сахар вызывает во рту ощущение песка. Белок, содержащийся в плодах западно–африканского растения Synsepalium dulcificum , изменяет кислый вкус на сладкий, так что лимон воспринимается как апельсин (см. Kurihara в ). Нанесение на язык кокаина вызывает последовательную утрату всех четырех типов вкусовых ощущений: горького, сладкого, соленого и, наконец, кислого.
Таблица 13.1. Классификация и характеристика химических чувств
|
Вкус |
Запах |
|
|
Рецепторы |
Вторичные сенсорные клетки |
Первичные сенсорные клетки; окончания |
|
Локализация рецепторов |
Язык |
V, IX и Х черепно–мозговых нервов, Нос и глотка |
|
Афферентные черепно–мозговые нервы |
VII, IX |
I, V, IX, Х |
|
Уровни синаптических переключении в ЦНС |
1.Продолговатый мозг 2.Вентральный таламус 3. Кора (постцентральная извилина) C вязи с гипоталамусом |
1. Обонятельная луковица 2.Конечный мозг (препириформная кора) Связи с лимбической системой и гипоталамусом |
|
Адекватные стимулы |
Молекулы органических и неорганических веществ, главным образом нелетучих. Источник стимулов – вблизи рецепторов или в непосредственном контакте с ними |
Почти исключительно молекулы органических летучих веществ в газовой фазе, растворяющиеся только вблизи рецептора. Источник стимулов, как правило, удален |
|
Число качественно различимых стимулов |
Мало (4 основных) |
Очень большое (тысячи) множество нечетко очерченных качеств |
|
Абсолютная чувствительность |
Сравнительно низкая (Не менее 10 16 молекул в 1 мл раствора) |
Очень высокая для некоторых веществ (10 7 молекул, у животных–до 10 2 –10 3 молекул в 1 мл воздуха) |
|
Контактное чувство. Используется для оценки качества пищи, регуляции ее потребления и переваривания (слюноотделительные рефлексы) |
Дистантное чувство. Используется для гигиенической оценки окружающей среды и пищи; у животных–для поисков пищи, коммуникации и в половом поведении. Включает сильный эмоциональный компонент |
У взрослых сенсорные вкусовые клетки расположены на поверхности языка. Вместе с опорными клетками они образуют в эпителии его сосочков группы из 40–60 элементов–вкусовые почки (рис. 13.1). Крупные, окруженные валиком сосочки в основании языка содержат до 200 вкусовых почек каждый; в более мелких грибовидных и листовидных сосочках на его передней и боковых поверхностях их лишь по нескольку. Всего у взрослого человека несколько тысяч вкусовых почек. Железы, расположенные между сосочками, выделяют омывающую вкусовые почки жидкость. Чувствительные к стимуляции дистальные части рецепторных (сенсорных) клеток образуют микроворсинки, выходящие в общую камеру, которая через пору на поверхности сосочка сообщается с внешней средой (рис. 13.1). Стимулирующие молекулы, диффундируя через эту пору, достигают вкусовых клеток (рецепторов).
Подобно прочим вторичным сенсорным клеткам, вкусовые генерируют при стимуляции рецепторный потенциал. Это возбуждение синаптически передается афферентным волокнам
Рис. 13.1. Схема вкусовой почки, погруженной в сосочек языка; показаны базальная, сенсорные, опорные клетки и афферентные волокна соответствующего черепномозгового нерва
черепно–мозговых нервов, которые проводят его в мозг в виде импульсов. В этом процессе участвуют: барабанная струна–ветвь лицевого нерва (VII), иннервирующая переднюю и боковые части языка, и языкоглоточный нерв (IX), иннервирующий его заднюю часть (рис. 13.2). Разветвляясь, каждое афферентное волокно получает сигналы от рецепторов разных вкусовых почек.

Рис. 13.2. Схема языка человека. Окраской выделена его иннервация различными черепно–мозговыми нервами; контурами обведены области распределения разных типов сосочков (1–грибовидные, 2–окруженные валиком, 3–листовидные). Локализация зон восприятия определенных вкусовых качеств показана значками
Замещаются вкусовые клетки очень быстро; продолжительность жизни каждой из них всего около 10 дней, после чего из базальной клетки формируется новый рецептор. Он устанавливает связь с афферентными волокнами таким образом, что их специфичность не меняется. Механизм, обеспечивающий такое взаимодействие, до сих пор не ясен (см. Oakley в ).
Реакции клеток в волокон . Одиночная вкусовая клетка в большинстве случаев реагирует на вещества, представляющие различные вкусовые качества, деполяризуясь или гиперполяризуясь ими (рис. 13.3). Амплитуда рецепторного потенциала повышается вместе с концентрацией стимулирующего вещества. На тип и амплитуду ответа влияет также окружающая среда (рис. 13.4).
Генераторный потенциал вызывает соответствующий уровень возбуждения афферентных волокон, формируя реакцию, называемую «вкусовым профилем» (рис. 13.5). Их импульсация зависит от реакции рецепторов следующим образом: деполяризация последних оказывает возбуждающее, гиперполяризация – тормозное действие.
Многие волокна IX пары черепно–мозговых нервов особенно сильно реагируют на вещества с горьким вкусом. Волокна VII пары сильнее возбуждаются при действии соленого, сладкого и кислого: некоторые из них сильнее реагируют на сахар, чем на соль, другие–на соль, чем на сахар, и т.д. Эти вкусоспецифичные особенности

Рис. 13.3. Внутриклеточные записи рецепторных потенциалов двух вкусовых клеток (а, б) языка крысы. Стимулы: 0,5 моль/л NaCI; 0,02 моль/л солянокислого хинина; 0,01 моль/л HCI и 0,5 моль/л сахарозы. Длительность действия каждого стимула показана горизонтальным отрезком (по Sato , Beidler в с изменениями)

Рис. 13.4. Влияние окружающей среды на форму и амплитуду внутриклеточных записей рецепторного потенциала одиночной вкусовой клетки языка крысы, стимулируемого 0,02 моль/л солянокислого хинина. Окружающая среда: а– 41,4 ммоль/л NaCI; б–дистиллированная вода (по Sato , Beidler в с изменениями)

Рис. 13.5. Ответы двух одиночных волокон барабанной струны крысы на различные вещества: 0,1 моль/л NaCI;
0,5 моль/л сахарозы; 0,01 н. HCI; 0,02 моль/л солянокислого хинина (по с изменениями)
разных групп афферентов обеспечивают информацию о вкусовом качестве, т.е. виде стимулирующей молекулы, а общий уровень возбуждения определенной их популяции–об интенсивности стимула, т. е. о концентрации данного вещества.
Центральные нейроны. Вкусовые волокна VII и IX пар черепно–мозговых нервов оканчиваются в пределах или в непосредственной близости от ядра одиночного пути (nucleus solitarius ) продолговатого мозга. Это ядро через медиальную петлю (медиальный лемниск) связано с таламусом в области его вентрального постеромедиального ядра. Аксоны нейронов третьего порядка проходят через внутреннюю капсулу и оканчиваются в постцентральной извилине коры головного мозга. В результате обработки информации на перечисленных уровнях число нейронов с высокоспецифичной вкусовой чувствительностью возрастает. Ряд корковых клеток реагируют только на вещества с одним вкусовым качеством. Расположение таких нейронов указывает на высокую степень пространственной организации вкусового чувства. Другие клетки в этих центрах реагируют не только на вкус, но и на температурную и механическую стимуляцию языка.
Вкусовая чувствительность человека
Вкусовые качества . Человек различает четыре основных вкусовых качества: сладкое, кислое, горькое и соленое, которые достаточно хорошо характеризуются типичными для них веществами (табл. 13.2). Вкус сладкого ассоциируется главным образом с природными углеводами типа сахарозы и глюкозы; хлорид натрия–соленый; другие соли, например KCI , воспринимаются как соленые и горькие одновременно. Такие смешанные ощущения характерны для многих естественных вкусовых стимулов и соответствуют природе их компонентов. Например, апельсин–кисло–сладкий, а грейпфрут
Таблица 13.2. Вещества с характерным вкусом и эффективность их воздействия на человека (Pfaffmann в )
|
Качество |
Вещество |
Порог восприятия, моль/л |
|
Горькое |
Сульфат хинина |
0,000008 |
|
Никотин |
0,000016 |
|
|
Кислое |
Соляная кислота |
0,0009 |
|
Лимонная кислота |
0,0023 |
|
|
Сладкое |
Сахароза |
0,01 |
|
Глюкоза |
0,08 |
|
|
Сахарин |
0,000023 |
|
|
Соленое |
NaCI |
0,01 |
|
CaCI 2 |
0,01 |
кисло–сладко–горький. У кислот вкус кислый; многие растительные алкалоиды горькие.
На поверхности языка можно выделить зоны специфической чувствительности. Горький вкус воспринимается главным образом основанием языка; другие вкусовые качества воздействуют на его боковые поверхности и кончик, причем эти зоны взаимоперекрываются (рис. 13.2).
Между химическими свойствами вещества и его вкусом не существует однозначной корреляции. Например, не только сахара, но и соли свинца сладкие, а самый сладкий вкус у искусственных заменителей сахара типа сахарина. Более того, воспринимаемое качество вещества зависит от его концентрации. Поваренная соль в низкой концентрации кажется сладкой и становится чисто соленой только при ее повышении. Чувствительность к горьким веществам существенно выше. Поскольку они часто ядовиты, эта их особенность предостерегает нас от опасности, даже если их концентрация в воде или пище очень низка. Сильные горькие раздражители легко вызывают рвоту или позывы на нее. Эмоциональные компоненты вкусовых ощущений широко варьируют в зависимости от состояния организма. Например, человек, испытывающий дефицит соли, считает вкус приемлемым, даже если ее концентрация в пище так высока, что нормальный человек от еды откажется.
Вкусовые ощущения, очевидно, весьма сходны у всех млекопитающих. Поведенческие эксперименты показали, что различные животные различают те же вкусовые качества, что и человек. Однако регистрация активности отдельных нервных волокон выявила и некоторые отсутствующие у человека способности. Например, у кошек обнаружены «водяные волокна», либо реагирующие только на раздражение водой, либо демонстрирующие вкусовой профиль, включающий воду в числе эффективных стимулов (см. Sato в ).
Биологическое значение . Биологическая роль вкусовых ощущений заключается не только в проверке съедобности пищи (см. выше); они также влияют на процесс пищеварения. Связи с вегетативными эфферентами позволяют вкусовым ощущениям влиять на секрецию пищеварительных желез, причем не только на ее интенсивность, но и на состав, в зависимости, например, от того, сладкие или соленые вещества преобладают в пище.
С возрастом способность к различению вкуса снижается. К этому же ведут потребление биологически активных веществ типа кофеина и интенсивное курение.
Поверхность слизистой носа увеличена за счет носовых раковин–гребней, выступающих с боков в просвет носовой полости. Обонятельная область, содержащая большинство сенсорных клеток,

Рис. 13.6. Схема полостей носоглотки человека (сагиттальный разрез). Обонятельная область ограничена верхней и средней раковинами. Показаны зоны, иннервируемые тройничным (V), языкоглоточным (IX) и блуждающим (X) нервами
ограничена здесь верхней носовой раковиной, хотя в средней также есть небольшие островки обонятельного эпителия (рис. 13.6).
Обонятельный рецептор–это первичная биполярная сенсорная клетка, от которой отходят два отростка: сверху – дендрит, несущий реснички, от основания аксон. Реснички, внутренняя структура которых иная, чем у обычных киноцилий, погружены в слой покрывающей обонятельный эпителий слизи и не способны активно двигаться. Пахучие вещества, приносимые вдыхаемым воздухом, вступают в контакт с их мембраной–наиболее вероятным местом взаимодействия между стимулирующей молекулой и рецептором. Аксоны, направляющиеся в обонятельную луковицу, объединены в пучки (fila olfactoria ). Во всей слизистой носа находятся, кроме того, свободные окончания тройничного нерва, и некоторые из них также реагируют на запахи. В глотке обонятельные стимулы способны возбуждать волокна языкоглоточного и блуждающего нервов (рис. 13.6). Слой слизи, покрывающий обонятельный эпителий, предохраняет его от высыхания и постоянно возобновляется за счет секреции и перераспределения киноцилиями.
Молекулы пахучих веществ поступают к рецепторам (сенсорным клеткам) периодически: во время вдоха ноздрями и в меньшей степени–из полости рта, диффундируя через хоаны. Таким образом, во время еды у нас возникают смешанные ощущения, в которых сочетаются вкус и запах пищи.
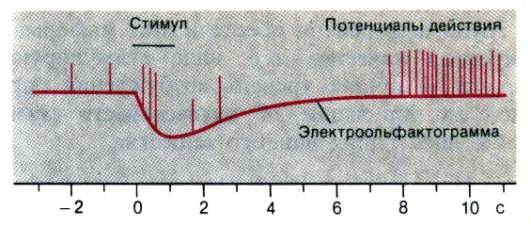
Рис. 13.7. Одновременная запись электроольфактограммы (красная линия) и потенциалов действия одиночного рецептора обонятельного эпителия лягушки при стимуляции нитробензолом. Длительность стимула (черный отрезок) –1 с (Gesteland в )
Обнюхивание–характерное поведение многих млекопитающих – значительно увеличивает приток воздуха к обонятельной слизистой и, следовательно, концентрацию в ней стимулирующих молекул.
Всего у человека в обонятельной области площадью примерно 10 см 2 около 10 7 рецепторов. У других позвоночных их бывает намного больше (у немецкой овчарки, например, 2,2–10 8). Обонятельные клетки, как и вкусовые, постоянно замещаются и из–за этого, по–видимому, не все функционируют одновременно.
Электроды, помещенные на обонятельный эпителий позвоночных, при действии запаха регистрируют медленные потенциалы сложной формы с амплитудой несколько милливольт. Эти электроольфактограммы (ЭОГ, рис. 13.7, см. Ottoson в ), как и электроретинограммы (ЭРГ), отражают суммарную активность многих клеток, поэтому не дают информации о свойствах индивидуальных рецепторов. Записать активность одиночного рецептора в обонятельной слизистой позвоночных удавалось только случайно (рис. 13.7). Показано, что спонтанная активность этих клеток очень низка (несколько импульсов в секунду), и каждая из них реагирует на множество веществ. Как и в случае вкусового профиля, можно построить спектр ответов одиночного обонятельного рецептора (см. Gesfeland в ).
Человек способен различать запах тысяч различных веществ. Обонятельные ощущения можно классифицировать на основе определенного их сходства, выявив определенные типы, или качества, запаха. Однако сделать это гораздо труднее, чем в случае вкусовых ощущений. Неопределенность категорий очевидна и в том, что классификации, предложенные разными авторами, неодинаковы. Корреляция между химической структурой и качеством запаха еще меньше, чем в случае вкусовых раздражителей. Табл. 13.3 показывает, что классы запахов, как правило, называют по их природным
Таблица 13.3. Отличительные характеристики классов запахов (Amoor , Skramlik )
|
Класс запахов |
Известные типичные вещества |
Сходство с запахом |
“Стандартный” источник |
|
Цветочный |
Гераниол |
Розы |
d –1–β–фенилэтилметилкарбинол |
|
Эфирный |
Бензилацетат |
Груши |
1,2–дихлорэтан |
|
Мускусный |
Мускон |
Мускуса |
3–метилциклопентадекан–1 –он |
|
Камфарный |
Цинеол, камфара |
Эвкалипта |
1,8–цинеол |
|
Гнилостный |
Сероводород |
Тухлых яиц |
Диметилсульфид |
|
Едкий |
Муравьиная кислота, уксусная кислота |
Уксуса |
Муравьиная кислота |
источникам или типичным веществам; каждую категорию можно также охарактеризовать «стандартным» источником.
Нейрофизиологические основы отнесения запахов к тому или иному классу до сих пор не обнаружены. Точка зрения, согласно которой группы, объединяющие близкие по запаху вещества, чем–то отличаются друг от друга, подтверждается случаями частичного нарушения обоняния (частичной аносмии). При таких дефектах (по крайней мере некоторые из них генетической природы) порог восприятия определенных обонятельных стимулов повышается. При этом часто он изменяется для нескольких веществ, относящихся, как правило, к одному и тому же классу запахов. Экспериментальные данные, применяемые для классификации запахов, можно получить при анализе перекрестной адаптации. Она заключается в том, что когда длительное действие какого–либо запаха вызывает повышение порога его восприятия, чувствительность к запаху некоторых других веществ также понижается (рис. 13.8). Изучая количественно такие взаимные изменения порогов, можно построить схему перекрестных адаптационных взаимоотношений (рис. 13.9). Однако и это не достаточно для однозначной и детальной классификации множества пахучих веществ по вызываемым ими ощущениям .
При интерпретации особенностей человеческого обоняния следует учитывать, что к пахучим веществам чувствительны и окончания тройничного нерва в слизистой носа, а также языкоглоточного и блуждающего нервов в глотке. Все они участвуют в формировании обонятельного ощущения (рис. 13.6). Их роль, никак не связанная с обонятельным нервом, сохраняется и при нарушениях функции обонятельного эпителия вследствие, например, инфекции (гриппа), опухолей (и связанных с ними операций на мозге) или черепно–мозговых травм. В подобных случаях, объединяемых термином гипосмия, порог восприятия существенно выше нормальною, однако способность различать запахи снижается лишь незначительно. При гипофизарном гипогонадизме (синдроме Кальмана) обоняние обеспечивается исключительно этими черепно–мозговыми нервами, поскольку при этом врожденном заболевании происходит аплазия обонятельных луковиц. Вредные температурные и химические воздействия могут вызвать обратимую или необратимую острую или хроническую гипосмию или аносмию в зависимости от природы и способа воздействия. Наконец, чувствительность к запахам снижается с возрастом.
Обоняние у человека очень чувствительно, хотя известно, что у некоторых животных этот аппарат еще более совершенен. В табл. 13.4 приведены концентрации двух пахучих веществ, достаточные для того, чтобы вызвать у человека соответствующие ощущения. При действии очень малых их количеств возникающее ощущение неспецифично; лишь после превышения некоторого порогового уровня запах не только выявляется, но и распознается. Например, скатол в низких концентрациях пахнет вполне приемлемо; при более высоких–отталкивающе. Таким образом, необходимо различать порог выявления и порог распознавания запаха.
Такие пороги, определяемые по ответам испытуемых или поведенческим реакциям животных, не позволяют установить чувствительность одиночной сенсорной клетки (рецептора). Однако, зная пространственную протяженность обонятельного органа человека и число рецепторов в его составе, можно вычислить и их чувствительность. Такие расчеты показывают, что одиночная сенсорная клетка деполяризуется и генерирует потенциал действия в ответ на одну, самое большее на несколько молекул пахучего вещества. Конечно, поведенческая реакция требует активации значительного числа рецепторов т.е. превышения определенного критического уровня отношения сигнал/шум в афферентных волокнах.
Кодирование. Кодирование обонятельных стимулов рецепторами пока можно описать только в первом приближении. Во–первых, индивидуальная сенсорная клетка способна реагировать на множество различных пахучих веществ. Во–вторых,

Рис. 13.8. Повышение интенсивности ощущения при увеличении интенсивности стимуляции (пропанолом) без адаптации (черная прямая) и после адаптации к пентанолу (красная кривая с черными треугольниками) (Cain , Engen в с изменениями)

Рис. 13.9. Перекрестные адаптационные взаимоотношения семи пахучих веществ: 1–цитраль, 2–циклопентанон, 3–бензилацетат, 4–сафрол, 5–м–ксилен, 6–метилсалицилат,7–бутилацетат.Реципрокные взаимодействия, как правило, неравносильны. Степень повышения порога восприятия обозначена следующим образом: черные линии–очень большая; красные сплошные линии большая; красные прерывистые линии–умеренная; красный пунктир–слабая (по с изменениями)
Таблица 13.4. Порог выявления для запахов масляной кислоты и бутилмеркаптана (Neuhaus , Stuiver )
|
Вещество г |
Число молекул в 1 мл воздуха |
Концентрация вещества вблизи источника стимула, моль/л |
|
Масляная кислота |
2,4–10 9 |
10 –10 |
|
Бутилмеркаптан |
10 7 |
2,7– 10 –12 |
у различных обонятельных рецепторов (как и у вкусовых) перекрывающиеся профили реакции. Таким образом, каждое пахучее вещество специфически возбуждает целую популяцию сенсорных клеток; при этом концентрация вещества отражается в общем уровне возбуждения.
Центральная обработка обонятельной информации
Обонятельная луковица . Гистологически обонятельная луковица подразделяется на несколько слоев, характеризующихся клетками специфической формы, от которых отходят отростки определенного типа с характерными связями между ними. Основные черты обработки информации здесь следующие: значительная конвергенция сенсорных клеток на митральных; выраженные тормозные механизмы и эфферентный контроль входной импульсации. В клубочковом (гломерулярном) слое аксоны приблизительно 1000 рецепторов оканчиваются на первичных дендритах одной митральной клетки (рис. 13.10). Эти дендриты образуют также реципрокные дендродендритные синапсы с перигломерулярными клетками. Митрально–перигломерулярные контакты–возбуждающие, противоположные по направлению–тормозные. Аксоны перигломерулярных клеток оканчиваются на дендритах митральных клеток соседнего клубочка. Такая организация позволяет модулировать локальный дендритный ответ, обеспечивая аутоторможение и торможение окружающих клеток. Клетки–зерна также образуют дендродендритные синапсы с митральными клетками (в данном случае–с их вторичными деидритами) и тем самым влияют на генерирование ими импульсов. Входы на митральных клетках тоже тормозные, т.е. реципрокные контакты участвуют в аутоторможении. Наконец, клетки–зерна образуют синапсы с коллатералями митральных клеток, а также с эфферентными (бульбопетальными) аксонами различного происхождения. Некоторые из этих центробежных волокон приходят из контралатеральной луковицы через переднюю комиссуру (спайку).
Особенность торможения, вызываемого лишенными аксонов клетками–зернами,–то, что в отличие от типичного торможения по Реншоу они могут активироваться частично, т. е. с пространственным градиентом. Эта

Рис. 13.10. Схема нейронных связей в обонятельной луковице. В клубочках (гломерулах) аксоны обонятельных рецепторов оканчиваются на первичных дендритах (D 1) митральных клеток. Перигломерулярные клетки и клетки–зерна образуют реципрокные синапсы с первичными и вторичными (D 2) дендритами митральных клеток. К–коллатерали. Направление синаптической передачи показано стрелками: возбуждающие влияния –черными, тормозные –красными (по с обобщениями и изменениями)
картина очень сложных взаимодействий вполне сравнима с известной в сетчатке, хотя обработка информации основана на другом принципе клеточной организации . Все описанное выше–лишь грубая схема происходящих в обонятельной луковице событий. Помимо митральных, к вторичным нейронам относятся и разнообразные пучковатые клетки, отличающиеся своими проекциями и медиаторами.
Центральные связи . Аксоны митральных клеток образуют латеральный обонятельный тракт, направляющийся в препириформную кору и пириформную долю. Синапсы с нейронами высших порядков обеспечивают связь с гиппокампом и, через миндалину, с вегетативными ядрами гипоталамуса. Нейроны, отвечающие на обонятельные стимулы, обнаружены также в ретикулярной формации среднего мозга и в орбитофронтальной коре.
Влияние обоняния на другие функциональные системы . Прямая связь с лимбической системой (см. разд. 16.6) объясняет выраженный эмоциональный компонент обонятельных ощущений. Запахи могут вызывать удовольствие или отвращение (гедонические компоненты), влияя соответствующим образом на аффективное состояние организма. Кроме того, нельзя недооценивать значение обонятельных стимулов в регуляции полового поведения, хотя результаты экспериментов на животных, особенно опытов по блокаде обоняния у грызунов, нельзя прямо переносить на человека. На животных также показано, что ответы нейронов обонятельного тракта можно изменить инъекцией тестостерона. Таким образом, на их возбуждение влияют и половые гормоны.
Функциональные нарушения . Помимо гипосмии и аносмии встречаются неправильное восприятие запаха (иаросмия) и обонятельные ощущения в отсутствие пахучих веществ (обонятельные галлюцинации). Причины таких расстройств различны. Например, они могут возникать при аллергических ринитах и травмах головы. Обонятельные галлюцинации неприятного характера (какосмия) наблюдаются главным образом при шизофрении.
Учебники и руководства
1. Beidler LM. (Ed.). Chemical Senses. Part 1. Olfaction. Part 2. Taste. Handbook of Sensory Physiology, Vol. IV, Berlin–Heidelberg–New York, Springer, 1971.
2. PfaffD. (Ed.). Taste. Olfaction and Central Nervous System. New York, Rockfeller University Press, 1985.
Оригинальные статьи и обзоры
3. Breipohl W. (Ed.). Olfaction and Endocrine Regulation, London, IRL Press, 1982.
4. Denton D. A., Coghlan J. P. (Eds.). Olfaction and Taste, Vol. V, New York, Academic Press, 1975.
5. Hayushi Т . (Ed.). Olfaction and Taste, Vol. II, Oxford–London New York–Paris, Pergamon Press, 1967.
6. Kare M. R., Mailer 0. (Eds.). The Chemical Senses and Nutrition, New York–San Francisco London, Academic Press, 1977.
7. Koster E. Adaptation and Cross Adaptation in Olfaction, Rotterdam, Bronder, 1971.
8. Le Magnen J., Mac Lead P. (Eds.). Olfaction and Taste., Vol. VI, London–Washington DC, IRL Press, 1977.
9. Norris D. M. (Ed.). Perception of Behavioral Chemicals, Amsterdam–New York–Oxford, Elsevier/North Holland, 1981.
10. Pfaffman С . (Ed.). Olfaction and Taste, Vol. Ill, New York, Rockfeller University Press, 1969.
11. Sato Т . Receptor potential in rat taste cells. In: Autrum H., Ottoson D., PerlE.R., Schmidt R.F., Shimazu H., Willis W.D. (Eds.). Progress in Sensory Physiology, Vol. 6, p. 1–37, Berlin–Heidelberg–New York–Tokyo, Springer, 1986.
12. Schneider D. (Ed.). Olfaction and Taste, Vol. IV, Stuttgart, Wiss, Verlagsges, 1972.
13. Shepherd G. M. Synaptic organization of the mammalian olfactory bulb. Physiol. Rev. 52, 864, (1972).
14. Van der Starre H. (Ed.). Olfaction and Taste. Vol. VII, London–Washington DC, IRL Press, 1980.
15. Zotterman Y. (Ed.). Olfaction and Taste. Vol. I. Oxford–London–New York Paris, Pergamon Press, 1963.
16. Chemical Senses. London. IRL Press (Published in regular installments).
Способность распознавать основные типы вкуса. Проверку сенсорной чувствительности по распознаванию основных типов вкуса проводят на модельных растворах химически чистых веществ:
Для приготовления растворов используют обработанную активным углем дистиллированную воду. Растворы хранят в колбах с притертой пробкой при температуре 18-20°С. Для опробования наливают в дегустационные бокалы по 35 мл раствора. Всего готовят девять проб: по 2 бокала с любыми тремя растворами и 3 бокала с четвертым раствором. Испытуемый не должен знать очередность подачи проб. Между пробами делают 1-2-минутный перерыв, ополаскивая обязательно рот чистой водой. При семи и более правильных ответах кандидата в дегустаторы рекомендуют для выполнения следующие тестовые задачи.
Определение индивидуального порога обнаружения вкуса.
С целью определения пороговой чувствительности к основным вкусовым ощущениям эксперту предлагают пробовать серию растворов возрастающей концентрации (табл.). Каждая серия состоит из 12 растворов.
Концентрация модельных растворов (в г/дм 3) для определения индивидуальных вкусовых порогов

Растворы готовят на обработанной активным углем дистиллированной воде. Концентрацию считают обнаруженной, если в трех треугольных сравнениях опознан исследуемый раствор. В каждой тройке растворов два являются растворителем, один - исследуемым. Подают их в возрастающей, в пределах одной тройки случайной, не известной испытуемому последовательности. Например:

Пороговая чувствительность к основным типам вкуса у кандидатов в органолептики- аналитики должна быть:
для сладкого вкуса < 7,0 г/дм по сахарозе;
для соленого вкуса < 1,5 г/дм по NaCl;
для кислого вкуса < 0,5 г/дм по винной кислоте;
для горького вкуса < 5,0 г/дм по MgS0 4 ;
Определение индивидуального порогового градиента вкусового восприятия. Пороговая разница вкусового восприятия представляет собой минимальную разность концентраций вкусового вещества, которую эксперт способен обнаружить. Определяется она в растворах с несильным, но хорошо опознаваемым вкусом. Абсолютное значение пороговой разницы зависит от концентраций раствора, поэтому чувствительность вкуса эксперта следует оценивать по индивидуальному
пороговому градиенту вкуса на модельных растворах (табл.). Например, 1 %-ный раствор глюкозы легко отличить от 2%-ного, по 20%-ный от 21%-ного - почти невозможно. В обоих растворах разность концентраций составляет 1%, но в первом случае градиент концентрации равен 2,0, во втором - 1,05.
Концентрация модельных растворов (в г/дм 3) для определения индивидуального порогового градиента вкуса

Для нахождения порогового градиента вкуса эксперту предлагают методом треугольных сравнений обнаружить опытную пробу на фоне растворов сравнения. Порядок подачи растворов такой же, как при определении порогов обнаружения основных типов вкуса.
Нулевые растворы представляют собой растворы сахарозы, хлористого натрия, винной кислоты и сернокислого магния фоновой концентрации. Пороговый градиент испытуемых должен быть:

Индивидуальный пороговый градиент (ИПГ) вкуса характеризует способность определять изменения во вкусе исследуемого продукта.
Оценка вкусовой памяти. Стабильность вкусовых и обонятельных ощущений (сенсорная память) является одним из наиболее ценных качеств дегустатора. Вкусовую память оценивают по способности определять интенсивность и качество
вкусовых ощущений. Кандидату в органолептики-аналитики предлагают пробовать 6-7 растворов и просят расставить их по возрастанию концентрации вкусового вещества. Аналогичная задача состоит в определении двух идентичных проб из семи растворов различной концентрации. Исследуемые растворы должны различаться больше, чем индивидуальный пороговый градиент вкуса оценщика. Градиент концентрации тестовых растворов находят по формуле 2ИПГ - 1. Например, если индивидуальный пороговый градиент оценщика 1,3, тестовый градиент будет равен 2x1,3- 1=1,6.
С целью определения стабильности вкусовых ощущений оценщик знакомится со вкусом растворов 8-10 веществ. Затем дают три пробы предыдущей серии, которые испытуемый должен опознать. Стандартную серию составляют из веществ, вкус которых имеет отношение к вину (в %): танин - 0,2, лимонная кислота - 0,5, уксусная кислота - 0,2, глюкоза - 2, янтарная кислота - 0,1, винная кислота -1, диэтиловый эфир винной кислоты - 0,2, молочная кислота - 0,03, сульфид натрия - 0,1. При тренировке вкусовой памяти тестовые задачи могут быть усложнены уменьшением градиента концентрации растворов и увеличением числа веществ, предлагаемых для опознания.
Роль вкусовой чувствительности в жизнедеятельности организма сводится, в первую очередь, к регуляции пищевого поведения (выбор определенного вида пищи, формирование реакций предпочтения и т.д.). Вкусовое восприятие не остается постоянным в течение длительного периода, оно меняется в зависимости от состояния организма и, в частности, от сиюминутной потребности в данном веществе. Можно сказать, что вкусовая система служит своего рода контрольным устройством, благодаря которому осуществляется выбор адекватных пищевых веществ.
Строение вкусовой системы
Рецепторы. Верхняя поверхность языка покрыта многочисленными складками слизистой оболочки, в толще которой расположены специализированные эпителиальные образования округлой формы. Это вкусовые почки или вкусовые луковицы. У человека в среднем насчитывается 9–10 тыс. луковиц. В толще луковицы имеется полость, сообщающаяся с внешней средой посредством отверстия – поры.
Каждая вкусовая луковица включает в себя от 30 до 80 уплощенных, вытянутых веретенообразных клеток, тесно прилегающих друг к другу наподобие долек апельсина (рис.10).
На поверхности каждой вкусовой клетки, обращенной в сторону поры, имеются микроворсинки, вступающие в контакт с растворенными веществами. Различают клетки трех типов – сенсорные, опорные и базальные, выполняющие разные функции. Сенсорные клетки являются зрелыми вкусовыми рецепторами, относящимися к типу вторичночувствующих. Базальные клетки – это незрелые рецепторы, они в процессе онтогенеза дают начало сенсорным клеткам. Опорные клетки выполняют вспомогательную функцию.
Проводящие пути системы вкусовой чувствительности. Каждая вкусовая клетка иннервируется одним, а чаще несколькими вкусовыми волокнами. Обнаружено, что с одной рецепторной клеткой могут образовывать синаптические контакты до 30 волокон.
В проведении сенсорной информации от вкусовых рецепторов принимают участие волокна лицевого, языкоглоточного, блуждающего и тройничного нервов. Основная и, по-видимому, наиболее специализированная информация связана с лицевым и языкоглоточным нервами.
От вкусовых луковиц передних двух третей языка отходят волокна, идущие в составе лицевого нерва. От задней трети языка, миндалин, твердого неба и глотки вкусовые импульсы поступают по языкоглоточному нерву.
Центральные отделы вкусовой системы. Все вкусовые волокна, вступающие в мозговой ствол, заканчиваются в ядре одиночного пучка, проходящего на всем протяжении продолговатого мозга. Это ядро является общим для лицевого, языкоглоточного и блуждающего нервов. От ядра одиночного пучка отходят волокна, которые направляются к дугообразному ядру, входящему в состав вентробазального комплекса таламуса. Оттуда информация передается к корковым центрам вкуса, главным образом, в нижнюю часть постцентральной извилины.
Физиология вкусовой системы
Одиночная вкусовая клетка в большинстве случаев реагирует на вещества разного вкусового качества, т.е. не является строго специфичной. Вместе с тем, порог чувствительности для одного из этих веществ обычно ниже, чем для других. Процесс взаимодействия молекулы вкусового стимула с рецептором на сегодняшний день до конца не выяснен. Большинство исследователей считают, что на мембране вкусовых клеток существуют специализированные активные центры, на которых осуществляется адсорбция молекулы раздражителя. Возможно, при взаимодействии рецептора с веществом происходит изменение конформации мембранных белков, что, в свою очередь, ведет к развитию рецепторного потенциала. Амплитуда рецепторного потенциала зависит от концентрации стимулирующего вещества.
Среди вкусовых волокон, проводящих информацию, были обнаружены волокна, избирательно реагирующие на раздражители определенного качества. С повышением концентрации раствора увеличивается как частота разрядов, так и число волокон, вовлекаемых в реакцию (за счет различных порогов). Однако большинство вкусовых волокон не являются строго специфичными. Так, многие волокна языкоглоточного нерва особенно сильно реагируют на вещества с горьким вкусом. А если учесть, что этот нерв иннервирует заднюю треть языка, то становится понятным, почему корень языка наиболее чувствителен к горькому. Это, однако, не означает, что к другим веществам языкоглоточный нерв нечувствителен. Его волокна могут реагировать и на другие вещества, если их концентрация будет достаточно высокой. Волокна лицевого нерва сильнее возбуждаются при действии соленого, сладкого или кислого: некоторые из них сильнее реагируют на сахар, чем на соль, другие – на соль, чем на сахар, и т.д. Таким образом, количество возбужденных волокон и уровень их возбуждения создают своеобразный узор импульсной активности, который и кодируют информацию о качестве и интенсивности раздражителя.
При рассмотрении физиологии центральных отделов вкусового анализатора необходимо отметить, что большая часть нейронов ядра одиночного тракта обнаруживают полимодальную чувствительность. Они реагирует не только на разные вкусовые качества, но также на тактильные и температурные раздражения поверхности языка. При этом каждая клетка имеет свой «профиль активности», т.е. определенный уровень реакции и рисунок ответа при действии того или иного вещества в определенной концентрации.
На более высоких уровнях сенсорной системы возрастает число нейронов с высокоспецифичной вкусовой чувствительностью. Так, при регистрации активности одиночных нейронов дугообразного ядра были обнаружены нейроны, специализированные для вкусовой, температурной и тактильной модальности. Ряд корковых клеток реагируют только на вещества с одним вкусовым качеством.
Основные характеристики вкусовой чувствительности
Основными характеристиками вкусовой чувствительности являются уровни абсолютных, дифференциальных порогов, латентные (скрытые) периоды вкусового ощущения и вкусовая адаптация.
Под абсолютным порогом понимают минимальную концентрацию химического вещества, вызывающую у человека вкусовое ощущение. Абсолютный порог восприятия разных вкусовых веществ может варьировать в значительных пределах (на несколько порядков). Как правило, пороги обнаружения горьких веществ значительно ниже, чем сладких, кислых и соленых. Это объясняется тем, что многие ядовитые соединения, опасные для здоровья и жизни организма, имеют горький вкус. Поэтому в процессе эволюции сформировался тонкий чувствительный механизм, способный предотвратить повреждающее действие таких веществ. Кроме того, порог может меняться для одного и того же вещества как у разных испытуемых, так и одного и того же индивидуума в разные периоды времени. Он зависит от места приложения и температуры тестирующего раствора, индивидуальных особенностей испытуемого, функционального состояния испытуемого в момент опыта, потребности организма в данном веществе. В связи со сложностью этой взаимосвязи полагают, что раздражение не является простым химическим взаимодействием вещества со вкусовой клеткой.
Разностный (дифференциальный) порог – это величина минимально ощутимой разницы в восприятии одного и того же вкусового раздражителя при переходе от одних концентраций к другим (выражается в единицах концентрации). Относительный дифференциальный порог есть отношение разностного порога к исходной концентрации (выражается в относительных единицах). Величина разностного и дифференциального порогов зависит от концентрации тестирующего раствора и места его приложения. Показано, что минимальный дифференциальный порог имеет место при средних концентрациях вещества, при низких и высоких концентрациях порог повышается.
Скрытый (латентный) период вкусовых ощущений – это время между нанесением раздражения и появлением ощущения вкуса. Латентный период уменьшается при увеличении концентрации тестирующего раствора и увеличении раздражаемой поверхности языка.
Явление адаптации заключается в повышении абсолютного порога и снижении интенсивности вкусового ощущения при длительном действии раздражителя. Зависит от вида и концентрации вещества. После прекращения действия раздражителя происходит восстановление чувствительности. Адаптация к сладким и соленым веществам происходит быстрее, чем к горьким и кислым.
Вопросы и задания для самоконтроля
1. Опишите строение вкусовой луковицы.
2. Что является причиной возникновения рецепторного потенциала во вкусовой клетке?
3. Как изменяется величина рецепторного потенциала при увеличении концентрации пахучего вещества?
4. Почему пороги обнаружения горьких веществ ниже, чем для других соединений?
Вкус - ощущение, возникающее при действии какого-либо вещества на вкусовые рецепторы языка и слизистой оболочки рта. В процессе эволюции вкус формировался как сенсорный механизм, способствующий выбору «хорошей» пищи, из чего следует, что вкусовые ощущения влияют на наши пищевые предпочтения. Кроме того, раздражение вкусовых рецепторов приводит к возникновению многочисленных врожденных (безусловных) рефлексов, управляющих деятельностью органов пищеварения. При этом в зависимости от свойств пищи секрет, выделяемый пищеварительными железами, может существенно изменять свой состав.
Вкусовые рецепторы - клетки, раздражение которых вызывает вкусовые ощущения. Большая часть их располагается на языке. Кроме того, вкусовые рецепторы расположены на задней стенке глотки, мягком небе и надгортаннике. Рецепторные клетки объединены во вкусовые почки (луковицы), а они собраны в три вида сосочков - грибовидные, желобовидные и листовидные.
Различные участки языка по-разному чувствительны к вкусовым модальностям. Основание языка, где преобладают желобовидные сосочки, наиболее чувствительны к горькому, кончик языка (на нем в основном грибовидные сосочки) - к сладкому, боковые части языка (листовидные сосочки) - к кислому и соленому.
Вкусовая почка лежит в толще многослойного эпителия. Она имеет форму луковицы и состоит из опорных, рецепторных и базальных клеток. В каждой почке несколько десятков клеток. Почки не достигают поверхности слизистой оболочки и связаны с ней через небольшие каналы - вкусовые поры. При этом рецепторные клетки образуют на своей вершине микроворсинки, находящиеся в общей камере непосредственно под порой. Вкусовые рецепторы - самые короткоживущие сенсорные клетки организма. Продолжительность жизни каждой из них около 10 дней, после чего так же, как и в случае обонятельной системы, из базальной клетки формируется новый рецептор. У взрослого человека 9-10 тысяч вкусовых почек. С возрастом часть их атрофируется.
Вкусовые рецепторы - вторичночувствующие. Сенсорные нейроны, которые проводят вкусовую информацию в ЦНС, - это псевдоуниполярные нейроны, входящие в состав ганглиев лицевого (VII пара), языкоглоточного (IX пара) и блуждающего (X пара) черепных нервов. Периферические отростки этих нейронов подходят к вкусовым рецепторам, и при достаточно сильном возбуждении рецепторов в ЦНС проводятся нервные импульсы. Вкусовые волокна оканчиваются на чувствительном ядре, расположенном в продолговатом мозге (ядро одиночного пути). Через это ядро поддерживается связь с безусловнорефлекторными центрами, осуществляющими простейшие рефлексы, например, слюноотделение, жевание, глотание. Горький вкус является сигналом для запуска ряда оборонительных реакций (выплевывание, рвота и т.п.).
Большая часть аксонов ядра одиночного пути перекрещивается, поднимается вверх к таламусу (где оканчивается на нейронах заднего вентрального ядра) и далее - к коре больших полушарий. В настоящее время выяснено, что вкусовые центры находятся в островковой доле коры, а также у нижнего конца центральной борозды (поле 43). Некоторое количество аксонов, идущих из продолговатого мозга, заканчивается в гипоталамусе. Они вносят свой вклад в управление уровнем пищевой и оборонительной мотиваций, генерацию положительных и отрицательных эмоций, а также определяют неосознанные пищевые предпочтения.
Выделяют пять основных вкусовых модальностей: сладкую, соленую, кислую, горькую и умами. Последняя модальность обозначается японским словом, обозначающим вкус глутамата натрия (хорошо выраженный мясной вкус). При изучении их особенностей применяют растворы различных веществ, которые капельно наносятся на разные участки языка. Как эталонное сладкое вещество применяют глюкозу, кислое - соляную кислоту, соленое - хлористый натрий (поваренная соль, NaCl), горькое - хинин. Каждая рецепторная клетка наиболее чувствительна к определенной вкусовой модальности, но отвечает и на другие виды вкусовой стимуляции (обычно значительно слабее, т.е. с более высоким порогом реакции).
«Сладкие», «горькие» и «умами» молекулы взаимодействуют с мембранными рецепторами, что в конечном итоге приводит к выбросу медиатора в синапсах между рецепторными клетками и волокнами сенсорных клеток и проведению нервных импульсов в ЦНС. Механизм генерации рецепторного потенциала при восприятии соленого и кислого вкуса отличается от обычного принципа работы хеморецепторов. В «соленых» рецепторных клетках присутствуют открытые натриевые каналы. Соленая пища содержит большое количество ионов Na + , поэтому он диффундирует (входит) внутрь вкусовых клеток, вызывая деполяризацию. Она в свою очередь приводит к выбросу медиатора. Кислый вкус вызывается большой концентрацией ионов водорода (Н +) в кислых продуктах. Входя в рецепторную клетку, они также вызывают деполяризацию.
Кроме вкусовых, в ротовой полости находятся также кожные рецепторы. В нормальных условиях целостное вкусовое восприятие формируется при их участии (определение консистенции пищи, ее температуры и т.п.). Более того, через тактильные рецепторы опосредуются такие, на первый взгляд, вкусовые ощущения, как ментоловое и жгучее (острое). В формирование вкусового восприятия вносит свой вклад и обонятельный анализатор. При нарушении обоняния (например, во время насморка) вкусовые ощущения значительно снижены.
Пороги чувствительности вкусовых рецепторов очень индивидуальны для разных людей (часть отличий генетически задана) и могут изменяться в зависимости от многих условий. Например, порог к хлористому натрию (поваренная соль) уменьшается при его удалении из пищи и увеличивается при беременности. Вкусовое ощущение зависит также от концентрации вещества. Так, максимально сладок 20% раствор сахара, максимально соленый 10% раствор хлористого натрия, максимально кислый 0,2% раствор соляной кислоты, максимально горький 0,1% раствор хинина. При дальнейшем повышении концентраций вкусовые ощущения уменьшаются. Зависят вкусовые ощущения и от температуры: «сладкие» рецепторы наиболее чувствительны при температуре пищи около 37С, «соленые» - примерно при 10С, при 0С вкусовые ощущения исчезают.
Как и все остальные сенсорные системы, вкусовая способна адаптироваться к постоянно действующему стимулу, и при длительном возбуждении рецепторов их порог увеличивается. Адаптация к одному из вкусовых ощущений часто снижает пороги для остальных. Это явление называется вкусовым контрастом. Например, после ополаскивания рта слабо подсоленным раствором чувствительность к другим вкусовым модальностям увеличивается.

 Как собрать и высушить листья смородины для чая?
Как собрать и высушить листья смородины для чая? К чему снится длинная дорога
К чему снится длинная дорога Обеспечение потребностей в ERP 1с торговля формирование заказов по потребностям
Обеспечение потребностей в ERP 1с торговля формирование заказов по потребностям Кто контролирует негосударственные пенсионные фонды
Кто контролирует негосударственные пенсионные фонды